El corazón, está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo.
Cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente

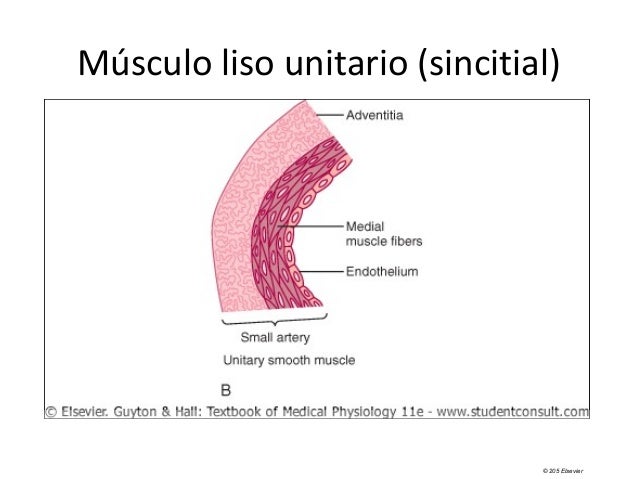
Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre: 1) hacia la circulación pulmonar por el ventrículo derecho, o 2) hacia la circulación sistémica por el ventrículo izquierdo. El músculo cardíaco es un sincitio de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí que cuando una célula se excita el potencial de acción se propaga rápidamente a todas.
Músculo cardíaco vs. Músculo esquelético.
- El potencial de acción del músculo esquelético está producido casi por completo por la apertura súbita de grandes números de canales rápidos de sodio que permiten que grandes cantidades de iones sodio entren en la fibra muscular esquelética desde el líquido extracelular. A estos canales se les denomina canales «rápidos» porque permanecen abiertos solo algunas milésimas de segundo y después se cierran súbitamente.
- En el músculo cardíaco, el potencial de acción está producido por la apertura de dos tipos de canales: 1) los mismos canales rápidos de sodio activados por el voltaje que en el músculo esquelético y 2) otra población totalmente distinta de canales de calcio de tipo L (canales lentos de calcio), que también se denominan canales de calcio-sodio.
- La segunda diferencia funcional importante entre el músculo cardíaco y el músculo esquelético que ayuda a explicar tanto el potencial de acción prolongado como su meseta es la siguiente: inmediatamente después del inicio del potencial de acción, la permeabilidad de la membrana del músculo cardíaco a los iones potasio disminuye aproximadamente cinco veces, un efecto que no aparece en el músculo esquelético.
Período refractario del músculo cardíaco.
- El período refractario normal del ventrículo es de 0,25 a 0,30 s, que es aproximadamente la duración del potencial de acción en meseta prolongado.
- Hay un período refractario relativo adicional de aproximadamente 0,05 s, durante el cual es más difícil de lo normal excitar el músculo pero, sin embargo, se puede excitar con una señal excitadora muy intensa, como se demuestra por la contracción «prematura» temprana del segundo.
- El período refractario del músculo auricular es mucho más corto que el de los ventrículos (aproximadamente 0,15 s para las aurículas, en comparación con 0,25 a 0,30 s para los ventrículos).

Duración de la contracción.
El músculo cardíaco comienza a contraerse algunos milisegundos después de la llegada del potencial de acción y sigue contrayéndose hasta algunos milisegundos después de que finalice. Por tanto, la duración de la contracción del músculo cardíaco depende principalmente de la duración del potencial de acción, incluyendo la meseta, aproximadamente 0,2 s en el músculo auricular y 0,3 s en el músculo ventricular.
CICLO CARDÍACO.

Los fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco hasta el comienzo del siguiente se denominan ciclo cardíaco.
- Cada ciclo es iniciado por la generación espontánea de un potencial de acción en el nódulo sinusal.
- NÓDULO SINUSAL: Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos.
Diástole y sístole.
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole.
La duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca.
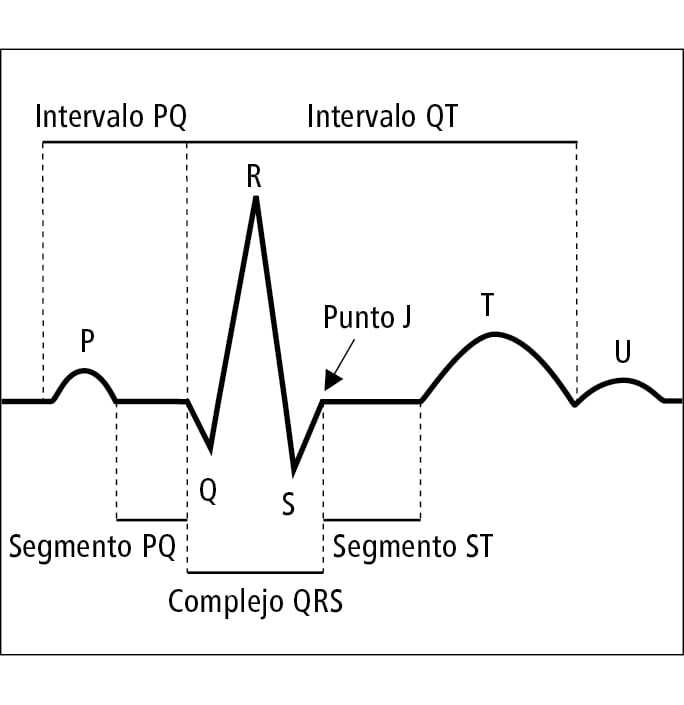
Las ondas P, Q, R, S y T, son los voltajes eléctricos que genera el corazón, y son registrados mediante el electrocardiógrafo desde la superficie del cuerpo.
- La onda P está producida por la propagación de la despolarización en las aurículas, y es seguida por la contracción auricular, que produce una ligera elevación de la curva de presión auricular inmediatamente después de la onda P electrocardiográfica.
- Aproximadamente 0,16 s después del inicio de la onda P, las ondas QRS aparecen comoconsecuencia de la despolarización eléctrica de los ventrículos, que inicia la contracción de los ventrículos y hace que comience a elevarse la presión ventricular.
- La onda T ventricular representa la fase de repolarización de los ventrículos, cuando las fibras del músculo ventricular comienzan a relajarse. Por tanto, la onda T se produce un poco antes del final de la contracción ventricular.
- Cambios de presión en las aurículas: las ondas a, c y v: En la curva de presión auricular se muestran tres pequeñas elevaciones de presión, denominadas curvas de presión auricular a, c y v.
- La onda a está producida por la contracción auricular. Habitualmente la presión auricular derecha aumenta de 4 a 6 mmHg durante la contracción auricular y la presión auricular izquierda aumenta aproximadamente de 7 a 8 mmHg.
- La onda c se produce cuando los ventrículos comienzan a contraerse; está producida en parte por un ligero flujo retrógrado de sangre hacia las aurículas al comienzo de la contracción ventricular, pero principalmente por la protrusión de las válvulas AV retrógradamente hacia las aurículas debido al aumento de presión de los ventrículos.
- La onda v se produce hacia el final de la contracción ventricular; se debe al flujo lento de sangre hacia las aurículas desde las venas mientras las válvulas AV están cerradas durante la contracción ventricular. Después, cuando ya ha finalizado la contracción ventricular, las válvulas AV se abren, y permiten que esta sangre auricular almacenada fluya rápidamente hacia los ventrículos, lo que hace que la onda v desaparezca.
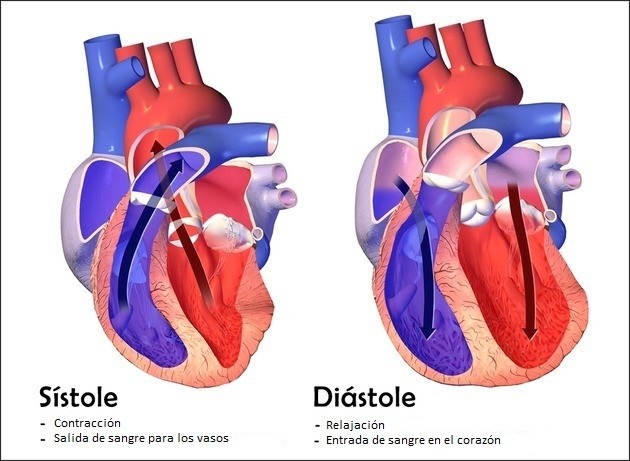
Los ventrículos se llenan de sangre durante la diástole.
Durante la sístole ventricular se acumulan grandes cantidades de sangre en las aurículas derecha e izquierda porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado la sístole y las presiones ventriculares disminuyen de nuevo a sus valores diastólicos bajos, el aumento moderado de presión que se ha generado en las aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y permite que la sangre fluya rápidamente hacia los ventrículos. Es el denominado período de llenado rápido de los ventrículos.
Período de relajación isovolumétrica (isométrica).
Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones intraventriculares derecha e izquierda disminuyan rápidamente. Las presiones elevadas de las grandes arterias distendidas que se acaban de llenar con la sangre que procede de los ventrículos que se han contraído empujan inmediatamente la sangre de nuevo hacia los ventrículos, lo que cierra súbitamente las válvulas aórtica y pulmonar. Durante otros 0,03 a 0,06 s el músculo cardíaco sigue relajándose, aun cuando no se modifica el volumen ventricular, dando lugar al período de relajación
isovolumétrica o isométrica. Durante este período las presiones intraventriculares disminuyen rápidamente y regresan a sus bajos valores diastólicos. Después se abren las válvulas AV para comenzar un nuevo ciclo de bombeo ventricular.
Volumen telediastólico, volumen telesistólico y volumen sistólico.
- Durante la diástole, el llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos hasta aproximadamente 110 a 120 ml. Este volumen se denomina volumen telediastólico.
- Después, a medida que los ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente 70 ml, lo que se denomina volumen sistólico.
- El volumen restante que queda en cada uno de los ventrículos, aproximadamente 40 a 50 ml, se denomina volumen telesistólico.
- La fracción del volumen telediastólico que es propulsada se denomina fracción de eyección, que habitualmente es igual a 0,6 (o el 60%) aproximadamente.

Válvulas auriculoventriculares.
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole.
Función de los músculos papilares.

Los músculos papilares que se unen a los velos de las válvulas AV mediante las cuerdas tendinosas. Los músculos papilares se contraen cuando se contraen las paredes ventriculares, pero, al contrario de lo que se podría esperar, no contribuyen al cierre de las válvulas. Por el contrario, tiran de los velos de las válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan demasiado hacia las aurículas durante la contracción ventricular.
Válvulas aórtica y de la arteria pulmonar.
Las válvulas semilunares aórtica y pulmonar funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas presiones de las arterias al final de la sístole hacen que las válvulas semilunares se cierren súbitamente, a diferencia del cierre mucho más suave de las válvulas AV. Segundo, debido a sus orificios más pequeños, la velocidad de la eyección de la sangre a través de las válvulas aórtica y pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores. Además, debido al cierre rápido y a la eyección rápida, los bordes de las válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas tendinosas, lo que no ocurre en el caso de las válvulas semilunares.
:max_bytes(150000):strip_icc():format(webp)/Valvulasdelcorazon-9ef5067bb6a14b2bacc0fb89ccd5f86a.jpg)
Regulación del bombeo cardíaco.
Cuando una persona está en reposo el corazón solo bombea de 4 a 6 l de sangre cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón bombee de cuatro a siete veces esta cantidad.
Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son: 1) regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón, y 2) control de la frecuencia cardíaca y del bombeo cardíaco por el sistema nervioso autónomo.
Control del corazón por los nervios simpáticos y parasimpáticos.
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón.
Para niveles dados de presión auricular de entrada, la cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática. Por el contrario, el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).